 RNA-verden
RNA-verden
(Fra boka 'The Design of Life' kap 8.6-7; av W. Dembski og J. Wells)
Det scenariet for opprinnelse til livet som har vakt mest oppmerksomhet siden 1990-tallet, er kjent under navnet 'RNA-verdenen'. Selv om eksperimenter ut fra simuleringer om primitiv atmosfære (lenke) kunne forklare dannelse av noen av livets elementære bestanddeler (aminosyrer), feilet de likevel i å forklare hvordan proteiner kunne dannes på den tidlige jorden. Og proteiner forutsetter DNA, og DNA forutsetter proteiner. Som en følge av dette, har 'opprinnelse-til-liv' forskere betraktet den muligheten at proteiner ikke var den første molekylære bygnings-stenen for livet. Da er ikke DNA noen god kandidat, fordi den trenger et helt kompleks av komplekse proteiner for å kopiere seg selv.
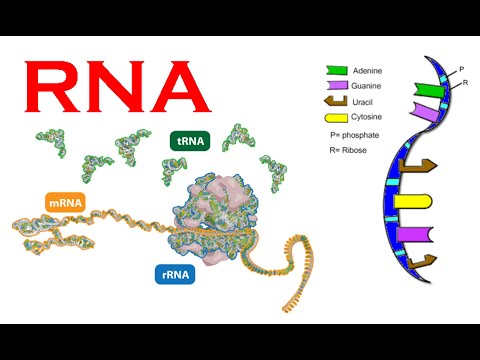
En annen kandidat er RNA. RNA er en nær kjemisk slektning av DNA, som benyttes av alle levende celler for å lage proteiner. På 1980-tallet viste molekylær-biolog Thomas Cech og Sidney Altman at RNA noen ganger kan oppføre seg som enzymer, det vil si som et protein som katalyserer kjemiske reaksjoner. Ut fra denne oppdagelsen foreslo molekylær biolog W. Gilbert at RNA kunne være i stand til å danne seg selv i fravær av proteiner. Denne RNA-verden kunne så ha vært det molekylære krybben som levende celler oppsto fra. RNA med katalytiske egenskaper, ble kjent som ribosymer, og søket etter slike RNA-forekomster: 'ribosym engineering'.
Bilde 1. Én RNA-verden hypotese
Men dette scenariet er også dypt problematisk. Ingen har vært i stand til overbevisende å demonstrere hvordan RNA kunne ha blitt formet før levende celler var der til å gjøre det. I følge biokjemiker Gerald Joyce (Scripps Research Institute) er ikke RNA en troverdig kandidat for livets første bygningsblokk, "fordi det er usannsynlig at de har blitt produsert i betydelige mengder på den tidlige jorda." (41) RNA består av nukleotid-baser (A,C,G eller U), ribose (sukkerart) og en fosfat-gruppe. Materielle prosesser som favoriserer dannelse av nukleotider, arbeider mot dannelse av korresponderende sukker eller fosfat-grupper, og vice versa. Faktisk har eksperimentell dannelse av RNA bare skjedd under de mest urealistiske prebiotiske forhold.
Ett slikt eks. på prebiotisk syntese ble publisert i Nature (1995), beskrevet av Robert Shaphiro: RNA-basen Cytosin ble forberedt i høy konsentrasjon ved å varme opp to rensede kjemikalier i en forseglet glasskolbe ved 100 grader C i omtrent en dag. Ét av reagerende stoffene (cyanocetaldehyd) er en reaktiv substans som er i stand til å reagere med et antall vanlige kjemikalier, som kan ha vært tilstede i den tidlige atmosfæren. Disse konkurrentene ble så ekskludert. Det var nødvendig med en ekstremt høy konsentrasjon for å få det andre deltakende stoffet, urea, til å reagere i tilstrekkelig grad for at reaksjonen skulle lykkes. Produktet cytocin kan selv-ødelegges ved enkel reaksjon med vann. . Denne eksepsjonelt høye konsentrasjonen ble sannsynliggjort i Nature ved å se for seg en visjon av en tørkende lagune på den tidlige jorda. Shaphiro svarte at han kalkulerte at lagunen måtte fordampe til størrelsen av en puddel, uten tap av innhold, for å oppnå den konsentrasjonen. Ikke noe lignende eksisterer på jorda i dag. (42)
 Bilde 2. Hvordan RNA bidrar ved protein-syntese
Bilde 2. Hvordan RNA bidrar ved protein-syntese
Det er heller ikke noe bevis for at noe slikt stoff eksisterte på den tidlige jorda. Men selv om RNA kunne dannes under realistiske prebiotiske forhold, ville det ikke eksistere lenge nok. Det er simpelthen for ustabilt. Forfatteren (Joyce) konkluderte derfor: "Det er mest sannsynlige tolkning at livet ikke startet med RNA." (43) Selv om han ennå tror at RNA verden gikk forut for DNA-verden, tror han at ett slags ikke-RNA liv gikk forut for RNA-liv igjen. I følge Joyce må man "bygge stråmann på stråmann for å komme til det punktet der RNA er et synlig første biomolekyl." (44)
RNA-verden scenariet tilbyr derfor ingen forklaring for hvordan et slikt "synlig første biomolekyl" kan ha oppstått. Men anta likevel at en hittil ukjent mekanisme kunne ha produsert et slikt molekyl. (45) Hvordan ville et slikt molekyl se ut, og hva ville det være i stand til å utvikle seg til? Men RNA-verden lar alle slike spørsmål stå ubesvart. RNA-verden forutsetter dermed et enda mer fundamentalt 'opprinnelse til liv' scenario.
Selv-organiserende verdener
Til tross for de store vanskelighetene med materialistiske forklaringer for livets opphav, er opprinnelse-til-liv forskere optimistiske m.h.t. utsikter for fremtidig suksess. Til tider kan en til og med få inntrykk av at bortsett fra noen få detaljer, så er problemet essensielt allerede løst. Som et eks. kan vi se på flg. bemerkning av matematikeren Ian Stewart: "Opprinnelse til livet synes ikke å være et spesielt vanskelig problem. Vi vet nå, i det minste på denne planeten, at nøkkel-ingrediensen er DNA. Livets basis er molekylært. Hva vi behøver er en forståelse av komplekse molekyler: Hvordan kan de ha oppstått, og hvordan de bidrar til den rike billedveven av levende former og oppførsel. .. Det er mange troverdige forklaringer; vanskeligheten er å velge mellom dem. Den overfloden reiser problemer for spørsmålet: "Hvordan begynte livet på jorda?" men ikke for det mer vesentlige emnet: "Kan liv oppstå fra ikke-levende prosesser?" (46)
Stuarts forsikring er for optimistisk. Vanligvis er det vanskelig nok å komme opp med én god teori for å gjøre rede for et fenomen. Faktisk innebærer påstanden om at det er mange troverdige forklaringer, at hver av dem blir mindre sannsynlig. Om én av dem virkelig var troverdig, ville vi kunne forvente en konsensus blant vitenskapsfolk om at den virkelig er sannsynlig (for ikke å uttale seg om den er en 'korrekt' forklaring.) I stedet finner vi en stor oppbygning av 'troverdige forklaringer', hvorav ingen hevdes universelt og der hver av dem har fatale mangler.
Når opprinnelse-til-liv forskere aksepterer troverdighet heller enn bevis som sin standard for vitenskapelig sannhet, så gir de i realiteten opp letingen etter det som virkelig hendte eller for det som med rimelig sannsynlighet kan ha hendt. Troverdighet medfører ikke noen anstrengelse for å estimere sannsynlighet, slik Stewart og andre benytter ordet. I stedet bestemmer de seg for hva de kan forestille seg var mulig eller kunne ha hendt. På denne måten erstatter de eksperimenter og data med meninger og fordommer. Hvor er vitenskapen i dette? På hvilken måte er troverdighet som Stewart tilskriver ulike materialistiske tilblivelsesmåter for livets opphav noe annet enn trosartikler?
Likevel stiller Stewart to spørsmål som fortjener nærmere oppmerksomhet: 1. Hvordan startet livet på jorda? og 2. Kan liv bli til av ikke-levende prosesser? Stewart ser på svaret på 1.spørsmål som under arbeid og det andre som et utvetydig ja. Men hva spør dette 2. spørsmålet om: Spør det bare om ikke-levende materie kan organiseres til materie slik den finnes i levende organismer, noe svaret er et klart ja på. All materie består av partikler som når de blir brutt langt nok ned, kan eksistere atskilt fra liv. Kosmologer har faktisk fastsatt at alle materie på jorda {-lenke} en gang fantes i stjerner, og aktive stjerner er for varme til å tillate liv.

Ved å si at liv kan dannes fra ikke-levende prosesser, sier dermed Stewart mer enn at ikke-levende materie kan arrangeres for å danne levende materie. Heller sier han at ikke-levende materie, uten noen hjelp utenfra, har evne til å organisere seg selv til levende materie. Faktisk betegner denne referansen til livets framdukking, innen 'opprinnelse-til-liv' forskning, at evt. evne ved ikke-levende materie til å organisere seg til levende materie, ikke skyldes noen vilt usannsynlig begivenhet, men er en iboende trekk ved ikke-levende materie. I den sammenheng skriver 'opprinnelse-til-liv' forsker H. Morowitz i boka 'The Emergence of Everything': synet på fremdukking av biokjemi som vi har diskutert, representerer et paradigme-skift fra hva leseren tidligere har møtt i biologi-kurs, der det ble formodet at tilfeldige produkt av fri radikale reaksjoner førte til monomerer, så til polymerer, så til celler (jfr. RNA-verden) I synet som utarbeides her, fører seleksjon til en kjerne metabolisme, som så produserer et ordnet hierarki av fremdukkende strukturer og funksjoner. Disse blir gradvis mer komplekse, og fører til den sofistikerte kjemien til den universelle stamfaren. Dette er et meget annerledes syn enn det som er blitt lært i standard introduksjonskurs, men jeg tror det er et mye mer troverdig scenario. (48)
Bilde 3. Metabolisme først-verden
Morowitz kontrasterer her sin 'metabolisme-først' tilnærming med den mer konvensjonelle 'genetikk-først' tilnærming, som betrakter nøkkelen til livets opprinnelse som dannelse av biologisk funksjonelle polymerer (som i RNA-verdenen). Ved overgang fra momomerer til polymerer, måtte det skje noen svært usannsynlige begivenheter. (49) Det skyldes at funksjonelle polymerer (om de ikke er ekstremt korte), finnes å være sjeldne blant totalen av mulige polymerer. Videre utøver ikke de fysiske egenskapene ved momomerer noen preferanse til en sekvens framfor en annen, i forhold til deres lineære arrangement.
F.eks. med RNA er sukker-fosfat grunnvollen (backbone) som nukleotidene fester seg til, komplett 'nøytral' til hvordan disse basene ordnes. Evnen polymerer har til å bære informasjon, skyldes det store antall frihetsgrader som fysikkens og kjemiens lover tillater at monomerer kan kombineres på. Disse lovene foretrekker ingen sekvenser framfor andre. Nettopp p.g.a. denne friheten er det ingen måte monomerer kan arrangere seg spontant på til biologisk funksjonelle polymerer, unntatt som vidstrakt usannsynligheter (unntatt da når de er svært korte). Dersom livets opprinnelse skyldes en ekstremt usannsynlig begivenhet, så utgjør ikke det noen vitenskapelig teori. En vitenskapelig teori om livets opprinnelse, er bare mulig om livet oppsto gjennom en serie steg der hvert steg er sannsynlig og sekvensen i det hele også er sannsynlig. I det tilfellet har ikke-levende materie en iboende evne til å organisere seg til levende. Det utgjør i så fall en selv-organiserende verden. Morowitz metabolisme-først tilnærming, er en forekomst av en selv-organiserende verden.
Foruten Morowitz selv-organiserende metabolisme-først verden, så har 'opprinnelse-til-liv' forskere foreslått mange andre selv-organiserende verdener. Vi tar med noen få (51):
G. Wächtershäuser så på bakterier som fortærer jern og svovel, har foreslått mineralet pyritt som en primær katalyst for livets opprinnelse. (54)
D. Deamer har foreslått at selv-organisering av lipider til to-lags vesikler (blærer), som ligner celle-membraner, var en kritisk faktor for livets opprinnelse. (56)

Men alle slike forslag er høyst spekulative, og adresserer bare ett eller noen få av de elementære aspektene ved livets opprinnelse, og er spinkle på detaljer jamvel i det de adresserer, har lite eksperimentell støtte og krever massiv intervensjon fra forskere for å oppnå interessante resultater (og feiler derved å reflektere realistisk prebiotiske forhold.)
F.eks. har H. Morowitz i årevis forsøkt å få prebiotiske kjemikalier til å organisere seg til en sitronsyre syklus. (59) Denne syklusen finnes i alle celler og utøver en form for selv-replikasjon i metabolske sykluser. Dette er et nøkkelsteg for Morowitz, men han kan ikke inntil i dag, oppnå dette uten ved hjelp av enzymer (60), som forutsetter livet -slik vi kjenner det, og dermed faller utenom 'opprinnelse-til-liv' forskningen.
D. Deamers har i sin lipid-verden demonstrert eksperimentelt at karbon-baserte molekyler, fra Murchisons meteoritt (som representerer en prebiotisk omgivelse), spontant organiserte seg selv til små kuler av mikrobe-størrelse. I tillegg besto kulene av to-lags lipider som finnes i celle-membraner. I følge R. Hazen er vesiklene som produseres i Deamers arbeid, "milevis unna aktuelle celle-membraner," som på sin side innehar ett nettverk av forbløffende ganger av protein-reseptorer som regulerer flyten av molekyler og kjemisk energi inn og ut av cellen." (61) Mange andre brikker i livets puslespill måtte falle på plass, om vi skulle hevde med visshet at Deamers lipid-verden spilte en viktig rolle i livets opprinnelse.
Bilde 4. 'Lipid-verden'
Om Wächtershäuser jern-svovel verden skulle lykkes, burde den kunne forklare hvordan en metabolsk sirkel kunne organisere seg. Det ville måtte ha en form for belegg på et mineral. Men hvorfor skulle det spille en avgjørende rolle i livets utvikling? Vi har ingen avgjørende grunn til å trekke en slik konklusjon, i mangel av en detaljert evolusjonær sti fra et slikt belegg til cellulært liv. men ingen slike stier er kjente. (Ikke engang stier som fullender den mye enklere RNA-verden er kjent.)
Å tro at cellulært liv oppsto i noen av disse verdener' er i fravær av harde bevis, ønsketenkning. Og slike bevis mangler notorisk. Uten å ta hensyn til det, fortsetter 'opprinnelse-til-liv' å betrakte selv-organisering som høyst relevant for livets opprinnelse. Se mer under molekylær darwinisme-lenke.
Referanser:
41. G. Joyce, "Evolution and the Origins of Life", Nature 338 (1989): 217-224
42. Shaphiro, "A Simpler Origin of Life,"
43. . G. Joyce, "Evolution and the Origins of Life", Nature 338 (1989)
44. Sitert i R. Irion, "RNA Can't Take the Heat," Science 279 (1998): 1303
45. Shaphiro, "A Simpler Origin of Life,"
46. Ian Stewart, Lifes Other Secret: The New Mathematics of the Living World (New York:John Wiley, 1998), 48
48. H. J. Morowitz, The Emergence of Everyting: How the World became Complex (New York: Oxford University Press, 2002), 76
49. Ca. 2/3 av forskere som publiserte innen opprinnelse-til-liv forskning støtter enda ideen at livet startet med den spontane dannelse av RNA eller en relatert selv-kopierende molekyl. Shaphiro refererer til de skremmende sannsynlighets-barrierene en slik forskning først må overkomme.
51. Eksistensen av den store variasjon av forklaringer, viser at opprinnelse-til-liv forskning ikke lar seg strupe av mangel på data. Fantasi kan være en god ting, men det er som å koke suppe på en spiker i forhold til vitenskap.
54. G. Wächtershäuser, "Evolution of the First Metabolic Cycles," Proceedings of the National Academy of Sciences 87 (1990): 200-204 og G. Wächtershäuser, "Life as We Don't Know It"," Science 289 (2000): 1307-1308
56. D.W. Deamer, "The First Living Systems: A Bioenergetic Perspective," Microbiology and Molecular Biology Reviews 61 (1997): 239-261
59. H. J. Morowitz, The Emergence of Everyting: How the World became Complex (New York: Oxford University Press, 2002),80
60. Hazen, Genesis, 209-210
61.
Hazen, Genesis, 151
Oversettelse og bilder ved Asbjørn E. Lund
{kind=link}
{kind=link}
{kind=link}